
выделите соответствующий фрагмент текста и нажмите Ctrl+Enter.

|
выделите соответствующий фрагмент текста и нажмите Ctrl+Enter. |
|
Бюллетень московского общества испытателей природы.
Отдел биологии, т. LXXII (1), 1967.
[114] — начало страницы.
OCR Bewerr, Halgar Fenrirsson
[114]
Домашние животные многочисленных кочевых племен, населявших в I и II тысячелетиях нашей эры обширные пространства степей и отчасти лесостепей юго-восточной Европы, до сих пор еще совершенно не изучены. И монографисты, и авторы, касавшиеся более частных вопросов истории всех этих племен, обращаясь к описанию животноводства, являвшегося основой хозяйственной деятельности и определявшего характер их жизненного уклада, ограничиваются показаниями древних арабских литературных источников и хорошо известными сообщениями Плано Карпини и Рубрука. «Они очень богаты скотом: верблюдами, быками, овцами, козами и лошадьми», — писал Плано Карпини о кочевниках Кыпчакской степи, простиравшейся от Днепра и на восток далеко за Волгу. Эти слова совершенно ясно характеризуют видовой состав домашних животных, но не дают представления об их физическом облике, породном составе и том значении, которое имели отдельные виды в общем составе стада. В настоящее время археологические данные позволяют несколько заполнить пробел в наших знаниях по всем этим вопросам.
В XIII в. нашей эры, в результате монгольских завоеваний возникло огромное феодальное государство Золотая Орда. В Восточной Европе в Золотую Орду входили обширные пространства среднего и нижнего Поволжья, половецкие степи, Крым и Северный Кавказ. Столицей этого государства был город Сарай Берке (ныне известный также под названием городища Царев), расположенный на берегу р. Ахтубы, неподалеку от г. Волгограда, где в настоящее время находится г. Ленинск. Раскопки этого замечательного памятника производились Г. А. Федоровым-Давыдовым в 1959—1963 гг. и дали, в частности, большое количество костных остатков животных, главным образом происходящих из слоя XIV в. нашей эры, результатам изучения которых посвящена настоящая статья. Татары в Золотой Орде были только привилегированным господствующим слоем, тогда как основное население составляли половцы (Греков, Якубовский, 1950; Федоров-Давыдов, 1966). Поэтому материалы, полученные при раскопках Сарая Берке, являются основой для изучения домашних животных кочевников юго-восточной Европы. [115]
В общей сложности из раскопок Сарая Берке было определено и исследовано более 24 тыс. костей (табл. 1), происходящих от 14 различных видов домашних и диких животных. Обнаруженный видовой состав не отличается, таким образом, разнообразием. Особенно это относится к диким животным, представленным в иследованных остатках всего 6 видами.
Видовой состав костных остатков
|
Вид |
количество костей |
минимальное количество особей |
|
Крупный рогатый скот |
8 916 |
136 |
|
Овца |
11 783 |
388 |
|
Свинья |
3 |
3 |
|
Лошадь |
3 134 |
69 |
|
Верблюд |
79 |
9 |
|
Собака |
412 |
25 |
|
Кошка |
4 |
1 |
|
Косуля |
2 |
2 |
|
Сайгак |
1 |
1 |
|
Кулан |
1 |
1 |
|
Лисица |
6 |
2 |
|
Перевязка |
2 |
1 |
|
Заяц-русак |
33 |
|
|
Всего |
24 376 |
645 |
Найденные в Сарае Берке дикие животные не вносят ничего принципиально нового в наши современные представления о фауне млекопитающих Нижнего Поволжья: сайгак, лисица, перевязка, заяц-русак и поныне здесь совершенно обычны. Хорошо известно и документально доказано изучением костных остатков из раскопок археологических памятников, что кулан был ранее широко распространен в степях юго-восточной Европы. Что касается косули, то, отмечая отсутствие точных сведений о прошлом распространении ее в Прикаспийской области, В. Г. Гептнер (Гептнер и др., 1961) проводит южную границу ареала этого вида на широте Волгограда. Находка костей косули в культурном слое Сарая Берке подтверждает справедливость этой точки зрения.
Обращает на себя внимание не только узость видового состава диких животных, но и малочисленность костных остатков их в Сарае Берке. Они составляют менее 0,2% от общего количества костей из этого археологического памятника и 2,2% минимального количества особей, которым они принадлежали. При этом большая часть остатков диких животных происходит от зайцев. Представляется достаточно очевидным, таким образом, что в жизни населения столицы Золотой Орды охота не имела сколько-нибудь существенного хозяйственного значения. Это и понятно, если учесть, что Сарай Берке был крупным политическим, торговым и ремесленным центром страны.
Обратимся теперь к описанию домашних животных, которым принадлежит большинство найденных при раскопках Сарая Берке костных остатков.
Крупный рогатый скот
Крупному рогатому скоту принадлежит почти 9 тыс. костей, что составляет несколько более 36,5% общего количества костных остатков животных, которые удалось определить из раскопок Сарая Берке. [116] Происходят они не менее как от 133 особей разного пола и возраста. По обоим этим показателям он уступает только мелкому рогатому скоту, занимая в остатках из золотоордынской столицы второе место. Многочисленные данные позволяют составить представление об индивидуальном возрасте животных, которым принадлежали кости из Сарая Берке. Так, например, изучение состояния системы настоящих коренных зубов нижних челюстей (130 экз.) обнаруживает следующее распределение их по возрасту животных (в %):
|
возраст от 5-6 до 15-18 месяцев (М1 имеется — М2 — отсутствует) |
4,6 |
|
возраст от 15-18 до 24-28 месяцев (М2 имеется — М3 отсутствует) |
19,2 |
|
возраст старше 24-28 месяцев (М3 имеется). |
76,2 |
|
|
100,0 |
Как показывают приведенные цифры, более 3/4 всех исследованных нижних челюстей происходит от животных старше 24-28 месяцев, т. е. уже взрослых.
К аналогичным результатам приводит изучение еще более многочисленных данных по состоянию синостозов у костей конечностей. Известно, в частности, что процесс срастания нижних эпифизов метаподий с телом кости завершается к 24-28 месяцам (Zietzschmann, Krölling, 1955; Habermehl, 1961). У изученных с этой целью 285 экз. пястных и плюсневых костей только 12% оказались еще свободными от эпифизов и, следовательно, 88% принадлежали животным старше 2 — 2 1/2 лет. Из 234 пяточных костей у 33% экз. еще не произошло срастание tuber calcanei с телом кости, которое наблюдается в 3-летнем возрасте, тогда как у остальных 67% процесс синостозирования уже завершен и они должны быть отнесены к животным старше 3 лет.
Все эти данные показывают, что в преобладающем большинстве кости скота происходят от взрослых особей и, таким образом, использование в пищу молодых животных не имело у жителей Сарая Берке широкого распространения.
Как это обычно свойственно кухонным остаткам, кости сильно разрушены, что связано с использованием мяса животных в пищу. Особенно сильно разрушены черепа, так что для краниологического изучения могут быть использованы только сохранившиеся костные стержни рогов и нижние челюсти.
Количество костных стержней рогов, найденных в остатках из раскопок Сарая Берке, довольно велико (133 экз.), и они составляют 1,49% от общего количества костей крупного рогатого скота. Цифры подобного значения характерны для остатков именно рогатых популяций (Цалкин, 1964), так что комолость, видимо, не имела сколько-нибудь широкого распространения среди золотоордынского скота.
Приводимые ниже биометрически обработанные данные измерений дают представление о размерах костных стержней рогов из Сарая Берке:
|
длина по большой кривизне (n = 18) |
156,60 ± 6,75 (111-220) мм |
|
обхват у основания (n = 53) |
142,80 ± 3,43 (101-195) мм |
|
индекс массивности (n = 18) |
87,20 ± 1,56 (72-100) % |
|
индекс уплощенности (n = 53) |
81,55 ± 0,86 (69-95) % |
Особенно крупных экземпляров среди имевшихся в нашем распоряжении не было. Примерно в таких же пределах варьируют по длине и в обхвате у основания одноименные кости у древнерусского лесного скота (Цалкин, 1956). В среднем, однако, величина их на 10-12 мм меньше, чем у золотоордынского. [117]
Гораздо более обширны данные по ряду костей посткраниального скелета. К сожалению, в силу уже упоминавшегося искусственного разрушения длинных трубчатых костей конечностей, обычно приходится ограничиваться измерениями ширины эпифизов их, что не дает представления об общих размерах той или иной части скелета. Исключение составляют только пяточные и таранные кости, фаланги пальцев, а также некоторые пясти и плюсны. Изучение сохранившихся целыми метаподий имеет особенно важное значение, поскольку оно открывает известные возможности судить как о составе популяции по полу животных, так и о росте скота (Цалкин, 1960).
Размеры метаподий скота
|
Признаки |
коровы |
быки |
||||
|
Lim |
M |
n |
Lim |
M |
n |
|
|
Длина пясти в мм |
176-209 |
192,50 |
21 |
187-194 |
192,2 |
3 |
|
Ширина верхнего конца в мм |
49-68,5 |
58,6 |
15 |
- |
62,5 |
1 |
|
Ширина нижнего конца в мм |
53-68,5 |
61,2 |
18 |
65,5-71 |
68,2 |
3 |
|
Ширина диафиза в мм |
27,5-37,5 |
33,1 |
21 |
38-39 |
38,5 |
2 |
|
Индекс ширины верхнего конца в % |
26,9-35,3 |
30,6 |
15 |
- |
32,2 |
1 |
|
Индекс ширины нижнего конца в % |
28,8-34,0 |
31 ,6 |
18 |
33,8-38,0 |
35,6 |
3 |
|
Индекс ширины диафиза в % |
15,1-18,6 |
17,4 |
21 |
19,6-20,1 |
19,9 |
2 |
|
Длина плюсны в мм |
203-224 |
219,0 |
5 |
218-225 |
221,5 |
2 |
|
Ширина верхнего конца в мм |
42-53,5 |
48,1 |
5 |
47,5-55 |
51,8 |
2 |
|
Ширина нижнего конца в мм |
56-65 |
58,5 |
5 |
60-65 |
62,5 |
2 |
|
Ширина диафиза в мм |
24,5-29,5 |
26,8 |
5 |
30,5-32 |
31,3 |
2 |
|
Индекс ширины верхнего конца в % |
19,1-23,9 |
22,0 |
5 |
21,8-24,4 |
23,1 |
2 |
|
Индекс ширины нижнего конца в % |
23,9-29,0 |
25,8 |
5 |
27,5-28,9 |
28,2 |
2 |
|
Индекс ширины диафиза в % |
11,0-13,2 |
12,3 |
5 |
14,0-14,2 |
14,1 |
2 |
Анализ абсолютных размеров и пропорций хорошо сохранившихся метаподий показывает (табл. 2, рис. 1 и 2), что большая часть их принадлежит коровам. Только 3 из 24 пястных и 2 из плюсневых костей, имеющие такую же длину, как и у коров, но отличающиеся большей массивностью, могут рассматриваться как принадлежащие быкам.
Таким образом, в исследованной серии метаподий, состоящей из 31 экземпляра, 26 или 84% приходится на долю коров и 5 или 16% — на долю быков. Если судить на основании этих данных о составе популяции скота в Сарае Берке, то должен быть особо отмечен высокий процент быков, значительно превышающий необходимый для обеспечения размножения.
Объяснение этому явлению можно видеть в использовании кочевниками быков в качестве транспортных животных, о котором сообщают многие древние литературные источники, в частности уже упоминавшиеся ранее Рубрук и Плано Карпини. Первый сообщает о том, что когда он собирался в путь из Солдайи (современный Судак) в куманские степи, ему были предложены для этой цели либо повозки, запряженные 2 волами каждая, либо вьючные лошади. Согласно Плано Карпини, кочевники перевозили свои войлочные жилища или на вьючных животных, или на повозках, в которые впрягались (в зависимости от величины жилища) от 1 до 4 и более волов. Для перевозки крупных жилищ использовались, по сведениям Рубрука, повозки еще больших размеров, в которые быки впрягались в два ряда, по два животных в каждом. Широкое использование быков (или волов) для транспортных целей, таким образом, не оставляет каких-либо сомнений. [118]
В связи с этим заслуживает внимания тот факт, что среди исследованных метаподий не было найдено экземпляров, которые можно было бы рассматривать как происходящие от волов. Приходится предположить, что кастрация быков производилась в сравнительно позднем возрасте, когда воздействие ее на рост метаподий уже не могло проявиться достаточно отчетливо.
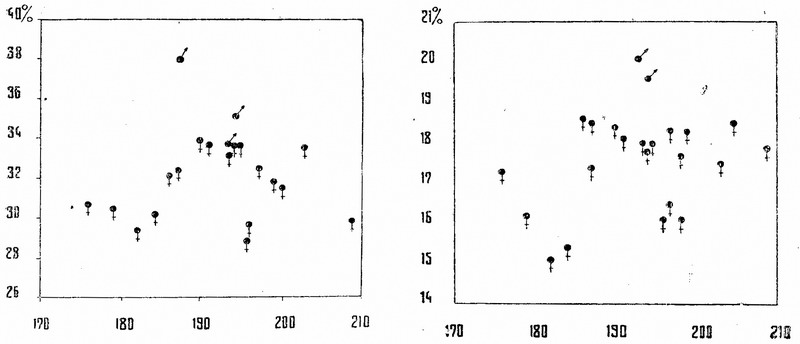
Рис. 1. График корреляции между длиной пясти и шириной нижнего конца кости крупного рогатого скота. На оси абсцисс — длина пясти в мм, на оси ординат — ширина нижнего конца пясти в % к общей длине кости
Рис. 2. График корреляции между длиной пясти и шириной диафиза кости крупного рогатого скота. На оси абсцисс — длина пясти в мм, на оси ординат — наименьшая ширина диафиза пясти в % к общей длине кости
Определяя на основании длины метаподий рост в холке у золотоордынского скота при помощи разработанных для этой цели коэффициентов (Цалкин, 1960), мы получаем следующие цифры:
коровы (n=26) 115,00±0,87 (105-125) см
быки (n=5) 121,0 (116-125) см
Насколько позволяют судить имеющиеся данные, рост золотоордынского скота варьировал в довольно широких пределах, от 105 до 125 см. У быков высота в холке была примерно на 6 см больше, чем у коров. В среднем рост рассматриваемого скота был 116,05 ± 0,86 см.
Нам остается теперь сравнить результаты измерений костей золотоордынского скота с аналогичными данными по другим группам древнего скота, разводившимся на территории Восточной Европы во II тысячелетии нашей эры. Мы имеем в виду в данном случае древнерусский лесной скот, разводившийся славянскими племенами в лесной полосе страны (Цалкин, 1956) и скот Волжских Болгар, известный по раскопкам г. Великий Болгар и других археологических памятников Среднего Поволжья (Цалкин, 1958). Для этой цели в табл. 3 приведены биометрически обработанные данные измерений некоторых костей скелета, часто встречающихся в остатках из раскопок.
Сравнивая между собой данные по древнерусскому лесному и золотоордынскому скоту, мы убеждаемся в существовании очень сильных различий. Они выражаются в диапазоне изменчивости, нижние и верхние границы которого у золотоордынского скота всегда выше, чем у древнерусского. Гораздо выше у золотоордынского скота значения [119]
Сравнение размеров костей крупного рогатого скота
|
Признаки |
древнерусский лесной |
болгарский |
золотоордынский |
M. diff |
||||
|
Lim |
M ± m |
Lim |
M ± m |
Lim |
M ± m |
древнерусский - золотоордынский |
золотоордынский - болгарский |
|
|
Длина альвеолярного ряда коренных зубов нижней челюсти в мм |
111-140 |
123,32 ± 0,30 |
130-146 |
137,50 ± 1,18 |
127-149 |
136,82 ± 1,45 |
9,1 |
0,4 |
|
Ширина нижнего суставного блока плечевой кости в мм |
58-77 |
63,76 ± 0,23 |
61-89 |
72,28 ± 1,12 |
61-84 |
74,16 ± 0,76 |
13,5 |
1,4 |
|
Ширина нижнего конца берцовой кости в мм |
45-67 |
52,88 ± 0,13 |
51-71 |
59,82 ± 0,68 |
51-68 |
60,90 ± 0,50 |
15,4 |
1,3 |
|
Длина пяточной кости в мм |
102-148 |
115,70 ± 0,31 |
115-151 |
128,48 ± 1,59 |
115-152 |
132,62 ± 1,07 |
15,2 |
2,2 |
|
Длина таранной кости в мм |
51-70 |
58,82 ± 0,10 |
57-73 |
62,88 ± 0,36 |
52-74 |
63,54 ± 0,37 |
12,4 |
1,3 |
|
Длина пясти в мм |
152-205 |
177,60 ± 0,32 |
172-215 |
188,95 ± 1,96 |
176-211 |
191,25 ± 1,05 |
12,5 |
1,0 |
|
Ширина нижнего конца пясти в мм |
45-70 |
50,48 ± 0,17 |
48-74 |
59,04 ± 0,95 |
49-74 |
61,92 ± 0,74 |
15,2 |
2,4 |
|
Индекс ширины нижнего конца пясти в % |
22,0-42,0 |
29,02 ± 0,10 |
26,0-36,0 |
31,74 ± 0,52 |
27,8-38 |
31,78 ± 0,46 |
5,7 |
0,1 |
|
Индекс ширины диафиза пясти в % |
12,0-22,0 |
15,40 ± 0,07 |
14,0-21 ,0 |
17,34 ± 0,36 |
15,1-20,1 |
17,67 ± 0,24 |
9,1 |
0,8 |
|
Длина больших фаланг в мм |
46-70 |
54,90 ± 0,10 |
52-74 |
60,45 ± 0,29 |
52-72 |
61,41 ± 0,17 |
34,3 |
2,8 |
[120] среднего арифметического всех исследованных признаков. Биометрический анализ устанавливает со всей очевидностью статистическую достоверность установленных различий (М. diff. для рассмотренных признаков составляет от 5,7 до 34,3). Интересно, что различия между древнерусским и золотоордынским скотом не ограничиваются абсолютными размерами костей, но проявляются и в их пропорциях, о чем свидетельствует сравнение относительной ширины нижнего эпифиза и диафиза пястей. Таким образом, следует признать, что кости золотоордынского скота не только крупнее, но и более массивны, чем у древнерусского. В то же время сравнение данных по скоту Волжских Болгар и Золотой Орды не обнаруживает различий по всем исследованным признакам.
Существенно различным оказывается и средний рост скота, что наглядно выражается следующими цифрами, характеризующими высоту в холке:
древнерусский лесной скот ...........106,30±0,16 (91-125) см
золотоордынский...............116,05±0,86 (105-125) см
Мы видим, что среди золотоордынского скота не встречаются такие очень малорослые животные, которые не только вполне обычны, но даже характерны в лесной полосе древней Руси. Нижние пределы изменчивости (рис. 3) у золотоордынского скота очень близки к средней величине признака у древнерусского, хотя среди последнего изредка наблюдаются животные, достигающие примерно тех же максимальных размеров, как и у золотоордынского. Если среди древнерусского скота для основной массы животных (88%) характерна высота в холке в пределах 100-115 см, то среди золотоордынского наиболее обычен рост 115-125 см. Особи высотой в холке более 115 см, составляющие только 4% у древнерусского лесного скота, образуют у золотоордынского наиболее многочисленную часть популяции. Средний рост скота Золотой Орды почти на 10 см превышает рост древнерусского. М. diff., вычисленный для рассматриваемого признака, составляет 11,1 и указывает на несомненную статистическую достоверность различий.
Итак, насколько можно судить на основании тех научных данных, которыми мы в настоящее время располагаем, во II тысячелетии нашей эры на территории Восточной Европы существовали две резко различные породы крупного рогатого скота, а именно:
1. Древнерусский лесной скот, малорослый и тонкокостный, разводившийся славянскими племенами в лесной полосе страны, а также племенами Восточной Прибалтики. Тот же или по крайней мере чрезвычайно близкий по размерам и структуре костей скот, судя по многочисленным публикациям зарубежных исследователей, встречался в Центральной, Западной и Северной Европе. К сожалению, до сих пор еще нет данных о скоте, разводившемся славянскими племенами в полосе лесостепи.
2. Степной скот кочевников юго-восточной Европы, гораздо более крупный и грубокостный, распространенный также и в Среднем Поволжье, у Волжских Болгар. По-видимому, в результате торговых связей скот этот проникает и на Верхнюю Волгу, где смешивается с древнерусским лесным. К подобному предположению заставляют прийти результаты изучения костных остатков из раскопок Старой Рязани.
Мелкий рогатый скот
Кости мелкого рогатого скота весьма многочисленны в культурном слое Сарая Берке и составляют почти 50% от общего количества [121] костных остатков животных, которые были найдены при раскопках этого памятника. Всего было определено 11783 кости, происходящие минимально от 388 овец и коз разного пола и возраста. В обоих этих отношениях, т. е. и по количеству костей, и по количеству особей, первое место среди животных из Сарая Берке принадлежит мелкому рогатому скоту.
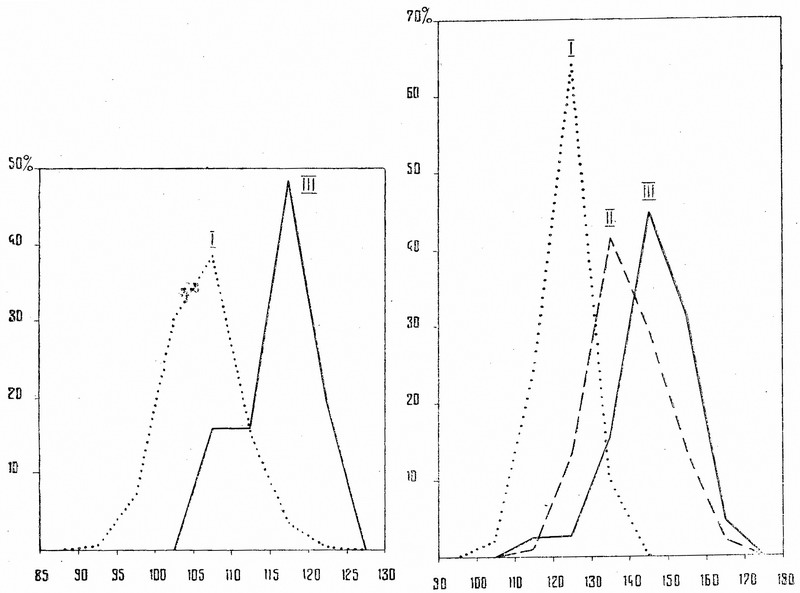
Рис. 3. Гистограмма распределения крупного рогатого скота по высоте в холке. На оси абсцисс — высота в холке в см, на оси ординат — количество экземпляров в %: I — древнерусский лесной скот; II — золотоордынский скот
Рис. 4. Гистограмма распределения пястных костей овец по общей длине. На оси абсцисс — длина пясти в мм, на оси ординат — количество экземпляров в %: I — древнерусская лесная овца; II — золото-ордынская овца; III — овца из раскопок Великого Болгара
Об индивидуальном возрасте животных, которым принадлежат рассматриваемые кости, позволяют судить следующие данные. Так, 543 изученные с этой целью нижние челюсти дают такую картину распределения по возрасту животных (в %):
|
возраст до 3 месяцев (М1 отсутствует) |
0,2 |
|
возраст от 3 до 9—12 месяцев (М1 имеется — М2 отсутствует) |
15,5 |
|
возраст от 9—12 до 18—24 месяцев (М2 имеется — М3 отсутствует) |
41,9 |
|
возраст старше 18—24 месяцев (М3 имеется) |
42,4 |
|
|
100,0 |
Мы видим, что большинство нижних челюстей (84,3%) принадлежит особям в возрасте от 9-12 до 18-24 месяцев и старше, причем обе эти возрастные группы представлены примерно равным процентом экземпляров. Следовательно, овцы и козы в более раннем возрасте употреблялись в пищу в ограниченном количестве. [122]
Еще более многочисленные данные для суждения о возрасте животных мы получаем при изучении синостозирования эпифизов костей конечностей, о сроках которого приводятся сведения в уже упоминавшихся нами сводках Цитшмана-Крёллинга и Хабермеля:
возраст до 3-4 месяцев (плечевая кость свободна от нижнего эпифиза).....................11,7 (n=408)
возраст моложе 15-20 месяцев (берцовая кость свободна от нижнего эпифиза.)...................31,7 (n = 458)
возраст моложе 20-24 месяцев (метаподии свободны от нижнего эпифиза).....................50,4 (n=375)
возраст моложе 3 лет (пяточная кость свободна от tuber calcanei).......................54,2 (n=296)
возраст моложе 3 - 3 1/2 лет (лучевые и бедренные кости свободны от нижних эпифизов)..............54,5 (n=180)
Приведенные выше цифры дают картину возрастного состава использованных в пищу животных аналогичную той, которая была получена при изучении нижних челюстей. Почти половина всех животных имела возраст 2-3 года и старше, 2/3 — старше 15-20 месяцев. Возрастная группа до 3-4-месячного возраста оказывается в данном случае представленной гораздо более высоким процентом особей, чем это было установлено при изучении нижних челюстей. В связи с этим надо заметить, что результаты определения численности этой последней группы по плечевым костям, вероятно, более соответствуют действительному положению вещей, потому что нижние челюсти очень молодых животных, тонкие и хрупкие, гораздо хуже сохраняются, чем челюсти взрослых.
Различение костных остатков овец и коз возможно далеко не по всем костям и трудности работы в этом направлении особенно усиливаются в связи с плохой сохранностью материала, связанной с использованием мяса животных в пищу. Тем не менее изучение количественных соотношений между двумя этими видами в остатках из раскопок Сарая Берке обнаруживает огромное преобладание овец над козами. Так, из 300 доступных видовой дифференциации фрагментов черепа 287 (95,7%) оказались принадлежащими овцам и только 13 (4,3%) — козам. Равным образом, из 219 метаподий 206 (94,1%) принадлежали овцам и 13 (5,9%) — козам. Совершенно очевидно, что козы составляли лишь весьма незначительную часть общего поголовья мелкого рогатого скота, разводившегося в Золотой Орде.
По причине плохой сохранности материала мы почти совершенно лишены возможности дать краниологическую характеристику золотоордынских овец и коз.
Костные стержни рогов баранов немногочисленны и все настолько сильно разрушены, что непригодны для измерений. Костных стержней рогов овец не обнаружено совершенно. Зато встречено большое количество фрагментов мозгового отдела черепа, у которых стержни рогов отсутствуют совершенно или представлены лишь небольшими шероховатыми возвышениями на поверхности лобных костей. Несомненно, что большую часть поголовья составляли комолые животные.
Весьма интересную часть костных остатков овец образуют более или менее хорошо сохранившиеся метаподии, которые представлены 45 пястными и 32 плюсневыми костями. Изучение их обнаруживает чрезвычайно широкую изменчивость. Так, среди найденных пястей имеются экземпляры длиной от 119 до 164 мм. Если первые довольно обычны для субфоссильных овец Восточной Европы, то вторые редко встречаются не только среди древних, но и современных пород. Весьма [123] изменчива и относительная ширина эпифизов и диафизов метаподии, среди которых наблюдаются и очень легкие, грацильные экземпляры и весьма грубые, массивные. При рассмотрении пястей и плюсен обнаруживается по этим причинам картина замечательного разнообразия.
Сравнивая между собой длину метаподий и относительную ширину их диафизов у древнерусских и золотоордынских овец, легко установить, что у последних они значительно длиннее и отличаются большей массивностью (табл. 4 и рис. 4, 5), причем различия по всем рассмотренным признакам статистически достоверны. Таким образом, овцы у кочевого населения юго-восточной Европы были более крупными и ширококостными, чем в лесной полосе страны.
Однако оказывается, что еще более крупными и ширококостными были овцы, разводившиеся населением Волжской Болгарии. Хотя длина метаподий варьирует у них примерно в тех же пределах, что и у золотоордынских овец, средняя величина их значительно больше и различия также статистически достоверны. Заслуживает внимания то, что среди метаподий болгарских овец часто встречаются экземпляры необычайно ширококостные. Индекс ширины диафиза пястей достигает у них 14,3% общей длины кости, а плюсны — 11,4%, что совершенно не наблюдается среди древнерусских и золотоордынских овец. Из всех известных в настоящее время овец, разводившихся во II тысячелетии нашей эры в Восточной Европе, болгарские не только наиболее крупные, но и самые ширококостные.
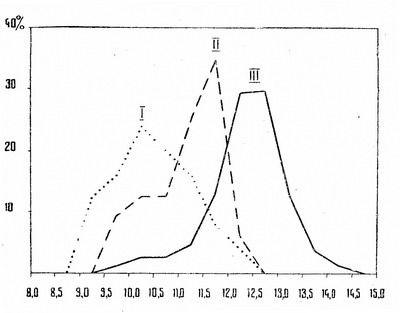
Рис. 5. Гистограмма распределения пястных костей овец по ширине диафиза. На оси абсцисс — ширина диафиза в % к общей длине кости, на оси ординат — количество экземпляров в %: I — древнерусская лесная овца; II — золотоордынская овца; III — овца из раскопок Великого Болгара
Определяя на основании общей длины метаподий рост в холке у золотоордынских овец по предложенным нами коэффициентам (Цалкин, 1961) устанавливаем, что он мог варьировать в пределах от 58 до 78, составляя в среднем 65-66 см.
Таблица 4.
Сравнение размеров метаподий овец*)
|
признаки |
древнерусская лесная |
золотордынская |
[124] болгарская |
M. diff |
||||
|
Lim |
M ± m |
Lim |
M ± m |
Lim |
M ± m |
Lim |
M ± m |
|
|
Длина пясти в мм |
110—135 |
123,20 ± 0,86 |
119—164 |
133,70 ± 1,41 |
118—161 |
140,40 ± 0,93 |
6,4 |
3,9 |
|
Индекс ширины диафиза пясти в % |
9,1—12,2 |
10,51 ± 0,11 |
9,6—12,3 |
11,16 ± 0,13 |
9,9—14,3 |
11,62 ± 0,08 |
3,8 |
3,7 |
|
Длина плюсны в мм |
121—150 |
132,35 ± 1,21 |
128—158 |
140,95 ± 1,41 |
131—169 |
148,30 ± 1,04 |
4,6 |
4,2 |
|
Индекс ширины диафиза плюсны в % |
7,5—9,8 |
8,73 ± 0,08 |
8,7—10,6 |
9,56 ± 0,11 |
8,7—11,4 |
9,73 ± 0,07 |
6,1 |
1,3 |
Изменчивость костей посткраниального скелета современных домашних овец сейчас еще очень мало изучена и это обстоятельство сильно затрудняет сравнение их с субфоссильными. По-видимому, по средней длине метаподий золотоордынские овцы близки к таким породам, как каракульская, романовская, северная короткохвостая, однако отличаются от них большей шириной диафиза (у упомянутых выше современных овец средняя ширина последнего составляет от 10,2 до 10,7% общей длины пястей).
Что касается коз, имевшихся у населения Сарая Берке, то кости их найдены, как уже упоминалось, только в небольшом количестве. Надо отметить, что все фрагменты мозгового отдела, принадлежащие как самцам, так и самкам, имеют костные стержни рогов. В отличие от овец, комолых экземпляров в остатках коз из раскопок столицы Золотой Орды не встречено.
Сохранившиеся метаподий представлены всего одной пястной и одной плюсневой костями, имеющими следующие размеры:
|
|
пясть |
плюсна |
|
длина кости в мм |
114 |
111 |
|
индекс ширины верхнего конца в % |
20,6 |
19,0 |
|
индекс ширины нижнего конца в % |
23,0 |
22,0 |
|
индекс ширины диафиза в % |
12,7 |
12,0 |
Одноименные кости коз подобных же размеров вполне обычны во II тысячелетии нашей эры в памятниках самых различных археологических культур Восточной Европы.
Свинья
Кости свиньи крайне редки в остатках из раскопок Золотой Орды. В 1960 г. были найден резец нижней челюсти взрослого животного и фрагмент верхней челюсти молодого в возрасте около шести месяцев. В 1962 г. была встречена еще одна кость свиньи — фрагмент нижней челюсти взрослой особи.
Как хорошо известно, кочевники юго-восточной Европы свиней вообще не разводили, поскольку это совершенно не совместимо с общим характером их жизненного уклада. Присутствие отмеченных выше немногих остатков свиней объясняется, надо полагать, тем, что в составе населения столицы Золотой Орды имелось известное количество русских ремесленников, которым и принадлежали эти животные. [125]
Лошадь
Кости лошадей вполне обычны в остатках из раскопок Сарая Берке, хотя далеко не так многочисленны, как кости крупного и мелкого рогатого скота. За все годы раскопок было определено 3134 кости, происходящие минимально от 69 особей.
Разрушение костей весьма сильно, а его характер не оставляет сомнений в том, что оно связано с использованием мяса лошадей в пищу. Столь же несомненно, что основная масса костей принадлежит взрослым животным. Резцы и коренные зубы молочной генерации хотя и попадаются, но в незначительном проценте. Довольно редки и свободные от эпифизов длинные трубчатые кости конечностей. Из 72 пяточных костей 77% имеют tuber calcanei уже сросшийся с телом кости и, следовательно, принадлежат особям старше трех лет. К подобным же выводам приводит и изучение синостозирования эпифизов длинных трубчатых костей. В пищу использовались, таким образом, преимущественно взрослые животные.
В Сарае Берке не было найдено ни целых черепов, ни крупных фрагментов их, которые давали бы возможность составить представление о краниологических особенностях золотоордынских лошадей; мало и хорошо сохранившихся длинных трубчатых костей конечностей. В дальнейшем обзоре мы остановимся более подробно только на некоторых костях посткраниального скелета, имеющих, как это принято считать, важнейшее значение для типологического изучения древних лошадей.
Наименьшая ширина диафиза пястей, выраженная в процентах от общей длины кости, в изученной серии из раскопок Сарая Берке варьирует от 14,0 до 16,5%. На основании величины рассматриваемого признака золотоордынские лошади распределяются следующим образом:
|
размеры в % |
золотоордынские |
древнерусские лесные |
|
крайнетонконогие (ширина диафиза менее 13,5) |
— |
3,7 |
|
тонконогие (ширина диафиза 13,6—14,5 |
25,0 |
25,6 |
|
полутонконогие (ширина диафиза 14,6—15,5) . |
50,0 |
48,2 |
|
средненогие (ширина диафиза 15,6—16,5) |
25,0 |
21,3 |
|
полутолстоногие (ширина диафиза 16,6—17,5) |
— |
1,2 |
Половину золотоордынских коней составляли, таким образом, полутонконогие, другую — примерно в равных количествах — тонконогие и средненогие. Нельзя не обратить внимания на то, что очень близкий характер имеет и распределение древнерусских лесных лошадей, среди которых наиболее многочисленную группу составляли также полутонконогие, в меньшей степени — тонконогие и средненогие. Правда, среди золотоордынских коней мы не находим ни крайнетонконогих, ни полутолстоногих, которые, хотя и в небольшом количестве встречались среди древнерусских, но это объясняется скорее всего просто ограниченностью изученной серии.
Путовые кости (первые фаланги пальцев) лошадей принадлежат к числу наиболее вариабильных элементов скелета, что проявляется как в общих размерах, так и в их пропорциях. Не составляют в этом отношении исключения и путовые кости золотоордынских лошадей. Лишь самые мелкие из них приближаются к размерам одноименных костей у [126] тарпана (Громова, 1949), но ни одна не достигает той замечательной массивности, которая считается столь типичной для этого последнего. В массе своей путовые кости лошадей Золотой Орды значительно крупнее и более стройные, чем у тарпана.
Очень сильная изменчивость отмечается и среди копытных костей (третьи фаланги пальцев), проявляющаяся при изучении всех размерных и структурных их признаков. Среди лошадей Золотой Орды были, несомненно, животные и с довольно узким, высоким копытом, и с очень широким и низким. Однообразия в этом отношении не было совершенно. Сравним между собой некоторые остеометрические данные по древнерусским и золотоордынским лошадям (табл. 5). По некоторым признакам (ширине нижнего суставного блока плечевой и лучевой кости,, ширине нижнего конца берцовой кости, длине пясти и передней путовой кости) между этими древними лошадьми удается обнаружить статистически достоверные различия. В других случаях различия подобного рода отсутствуют и характерно, что пропорции костей всегда очень близки. Таким образом, насколько позволяет судить исследованный материал, различия между древнерусскими и золотоордынскими лошадьми сводятся лишь к несколько более крупным абсолютным размерам костей у последних.
Серия хорошо сохранившихся длинных трубчатых костей конечностей дает возможность определить рост золотоордынских лошадей, пользуясь для этой цели данными В. О. Витта (1952).
|
|
Золотоордынские |
Древнерусские лесные |
|
Очень мелкие (высота в холке 112-120 см) |
— |
1,5 |
|
Мелкие (120-128 см) |
3,1 |
23,7 |
|
Малорослые (128-136 см). |
53,1 |
45,9 |
|
Средние (136-144 см) |
43,8 |
25,0 |
|
Рослые (144-152 см) |
— |
3,3 |
|
Крупные (152-160 см) |
— |
0,6 |
|
|
100% (n=32) |
100% (n=548) |
Степные лошади Восточной Европы в свете данных из раскопок Сарая Берке представляются довольно однородными по росту. Среди них не было обнаружено ни очень мелких, ни рослых и крупных, что, может быть, в известной степени находит себе объяснение в ограниченности количества наблюдений. Наиболее многочисленную группу среди них составляют малорослые лошади и немного меньшим процентом были представлены средние; мелкие были очень редки. При сравнении с аналогичными данными по древнерусским лошадям обращает на себя внимание заметное уменьшение у золотоордынских лошадей процента мелких и увеличение процента средних. Тенденция к увеличению роста выявляется, таким образом, с достаточной очевидностью. Что касается средней величины роста в холке, то у рассматриваемых субфоссильных лошадей она выражается следующими цифрами:
древнерусская лесная............132,56±0,28 см
золотоордынская...............135,28±0,76 см
В среднем высота в холке у золотоордынских лошадей почти на 3 см больше, чем у древнерусских, причем различия статистически достоверны (М. diff.-3,3).
Мы имеем, следовательно, возможность констатировать, что при отсутствии каких-либо существенных различий в структурных [127]
Таблица 5.
Сравнение размеров костей лошадей.
|
Признаки |
Древнерусская |
Золотоордынская |
M. diff |
||||
|
Lim |
M ± m |
n |
Lim |
M ± m |
n |
||
|
Ширина нижнего суставного блока плечевой кости в мм |
57—85 |
69,40 ± 0,42 |
145 |
67,5—76,5 |
71,37 ± 0,38 |
51 |
3,5 |
|
Ширина нижнего суставного блока лучевой кости в мм |
51—70 |
60,34 ± 0,23 |
238 |
60,5—68 |
63,34 ± 0,38 |
24 |
6,8 |
|
Ширина нижнего конца берцовой кости в мм |
58—80 |
69,36 ± 0,34 |
181 |
65—79,5 |
71,82 ± 0,60 |
34 |
3,6 |
|
Длина пясти в мм |
191—259 |
214,90 ± 0,86 |
165 |
207—236 |
221,25 ± 1,61 |
20 |
3,6 |
|
Индекс ширины диафиза пясти в % |
12,1—16,9 |
14,91 ± 0,06 |
164 |
14,0—16,5 |
15,00 ± 0,15 |
16 |
0,5 |
|
Длина плюсны в мм |
225—290 |
255,40 ± 0,98 |
155 |
240—280 |
259,8 |
5 |
— |
|
Индекс ширины диафиза плюсны в % |
10,2—13,7 |
11,69 ± 0,05 |
154 |
11,4—12,4 |
12,0 |
5 |
— |
|
Длина передней путовой в мм |
67—96 |
80,40 ± 0,49 |
132 |
76—92 |
83,60 ± 0,77 |
27 |
3,5 |
|
Индекс ширины диафиза передней путовой в % |
34,5—47,6 |
41,10 ± 0,19 |
131 |
37,6—45,2 |
41,86 ± 0,38 |
27 |
1,8 |
|
Длина задней путовой в мм |
65—98 |
77,80 ± 0,52 |
120 |
75-86 |
79,70 ± 0,51 |
34 |
2,6 |
|
Индекс ширины диафиза задней путовой в % |
37,5—45,7 |
41 ,34 ± 0,19 |
119 |
37,5—47,4 |
41,82 ± 0,32 |
34 |
1,3 |
|
Наибольшая ширина передней копытной кости в мм |
61—90 |
76,50 ± 0,73 |
48 |
69—84 |
77,72 ± 0,88 |
27 |
0,6 |
|
Индекс ширины передней копытной кости в % |
138,2—177,0 |
151 ,40 ± 1,34 |
48 |
139,2—171,4 |
152,30 ± 1,77 |
26 |
0,4 |
|
Индекс высоты передней копытной кости в % |
42,3—56,6 |
50,60 ± 0,48 |
48 |
44,6—54,2 |
50,24 ± 0,54 |
26 |
0,4 |
|
Наибольшая ширина задней копытной кости в мм |
64—87 |
74,70 ± 1 ,04 |
43 |
65—80 |
72,28 ± 0,66 |
27 |
2,0 |
|
Индекс ширины задней копытной кости в % |
127,0—154,9 |
138,30 ± 1,66 |
43 |
127,4—143,4 |
135,85 ± 0,89 |
27 |
1,3 |
|
Индекс высоты задней копытной кости в % |
49,3—60,6 |
54,55 ± 0,42 |
43 |
49,3—61,4 |
54,61 ± 0,67 |
27 |
0,1 |
[128] признаках посткраниального скелета, о которых позволяют судить изученные костные остатки, степные лошади Восточной Европы были несколько крупнее разводившихся в лесной полосе страны.
Верблюд
Кости верблюдов довольно редки в культурном слое Сарая Берке. За все годы раскопок было найдено всего 79 костей, принадлежащих минимально 9 особям. В их числе обломки черепа, нижние челюсти, позвонки, отдельные зубы, кости конечностей и их поясов.
Судя по характеру разрушения костей, мясо верблюдов, найденных в Сарае Берке, было использовано в пищу.
Собака
На общем фоне остатков животных кости собак выделяются своей гораздо лучшей сохранностью. Среди 412 определенных костей, происходящих минимально от 25 особей, имеются целые черепа и неповрежденные длинные трубчатые кости конечностей; последних значительно больше, чем разрушенных. Все эти признаки указывают на то, что мясо собак жителями города в пищу не употреблялось.
Кошка
Домашняя кошка уже была известна населению Сарая Берке, хотя, видимо, оставалась довольно редкой. В изученных остатках обнаружены только 4 кости (2 фрагмента нижней челюсти, лопатка и плечевая кость), принадлежащие одной особи.
Завершая на этом краткое описание остеологических особенностей домашних животных, обратимся теперь к анализу количественных соотношений между ними, позволяющему судить об удельном весе того или иного вида в составе стада и роли его в хозяйственной жизни населения.
Природные условия юго-восточной Европы (обширные пространства степей и полупустынь, незначительное количество осадков) способствовали развитию кочевого животноводства, связанного с круглогодичным содержанием скота на естественных пастбищах. Ценные продукты животноводства создавались при сравнительно небольших затратах труда и средств производства. Основное поголовье сельскохозяйственных животных составляли овцы, лучше других видов использующие пастбища и более полно удовлетворяющие потребности натурального хозяйства кочевников. На долю овец, в свете изучения остатков из раскопок Сарая Берке, приходилось почти 2/3 от общего поголовья сельскохозяйственных животных. Они разводились преимущественно для употребления в пищу и были, во всяком случае по своей численности, важнейшим объектом животноводства. Количество коз, как уже отмечалось выше, было весьма ограничено. Но в условиях кочевого хозяйства козы получают широкое распространение там, где возникает необходимость использовать бедные пастбища крутых горных склонов и растительность скалистых стаций, т. е. мест сравнительно мало пригодных для овец, не говоря уже об остальных видах сельскохозяйственных животных.
Вторым по численности видом, значительно уступавшим, однако, в этом отношении овцам, был крупный рогатый скот. Им широко [129] пользовались для молочных и транспортных целей, но он был также и важным источником мясной пищи. Третье место занимала по своей численности лошадь, использовавшаяся, видимо, наиболее разносторонне: под седлом, как вьючное и как упряжное животное (об этом свидетельствуют имеющиеся в Кенигсбергской летописи изображения половцев). Подобно крупному рогатому скоту, лошадь была источником получения молочных продуктов. В связи с этим заметим, что именно последние служили в летнее время основной пищей кочевников, тогда как мясо, согласно наблюдениям Рубрука, сушили и употребляли в пищу главным образом в зимние месяцы. Верблюды были, по-видимому, малочисленны и образовывали очень незначительную часть стада сельскохозяйственных животных. Ими пользовались как вьючными животными и, по свидетельству Рубрука, запрягали в повозки небольших размеров; употребление мяса верблюдов в пищу не имело широкого распространения. Очень характерно для кочевого животноводства отсутствие свиней; небольшое количество их, обнаруженное в остатках из раскопок Сарая Берке, связано с особенностями этнического состава населения золотоордынской столицы и может в данном случае не приниматься во внимание.
Таблица 6
Соотношения между видами сельскохозяйственных животных по количеству особей
|
Место находок |
Всего особей |
В том числе в % |
||||
|
Крупный рогатый скот |
Мелкий рогатый скот |
лошадь |
верблюд |
всего |
||
|
Великий Болгар (слой XIII— XIV вв.) |
1044 |
13,8 |
76,4 |
9,2 |
0,6 |
100,0 |
|
Сарай Берке (слой XIV в.) |
602 |
22,6 |
64,4 |
11,5 |
1,5 |
100,0 |
С той же в сущности картиной количественных соотношений между сельскохозяйственными животными сталкиваемся и при изучении костных остатков из раскопок Великого Болгара. Различия выражаются лишь в несколько более высоком проценте мелкого рогатого скота и более низком — всех остальных видов. По-видимому, хозяйство населения Великого Болгара находилось под сильным влиянием традиций кочевого животноводства. Возможно, что наблюдающееся уменьшение относительной численности крупного рогатого скота, лошадей и верблюдов отражает оседлость болгарского населения, уже не испытывавшего в силу этой причины столь высокой потребности в транспортных животных.
Институт археологии АН СССР, Москва
Summary
Bone remains of the animals found in the excavations of the town of Saray Berke (close to the town of Volgograd), the capital of the feudal state «Golden Horde» that had appeared in the XIII Century were taken under study. More than 24 thousands of bones belonging to 8 species of domestic and 6 species of wild mammals (Table I) were identified. The data obtained give the idea on the domestic animals and stock breeding [129] of the nomads of the south-eastern Europe that were the main population of the Golden Horde.
Of the farm animals of the Golden Horde population sheep were most numerous (there were but few goats), followed by cattle and then by horses; cameis were not numerous (Table 6).
The cattle and sheep of the nomads of south-east Europe differed distinctly from those bred by Slav tribes in considerably larger dimensions and massivity of their skeleton; the horses of nomads were also larger.
Витт В. О. 1952. Лошади Пазырыкских курганов. «Советская Археология», т. XVI.
Громова Вера. 1949. История лошадей в Старом Свете. «Тр. Палеонтологич. ин-та АН СССР», т. XVII. М., Изд-во АН СССР.
Гептнер В. Г., Насимович А. А., Банников А. Г. 1961. Млекопитающие Советского Союза, т. I. Парнокопытные и непарнокопытные. М., «Высшая школа».
Греков Б. Д., Якубовский А. Ю. 1950. Золотая Орда и ее падение. М.-Л. Изд-во АН СССР.
Карпини П. История монголов. В кн.: Путешествие в восточные страны Плано Карпини и Рубрука. М., Соцэкгиз, 1957.
Рубрук В. Путешествие в восточные страны. В кн.: Путешествие в восточные страны Плано Карпини и Рубрука. М., Соцэкгиз, 1957.
Федоров-Давыдов Г. А. 1966. Кочевники Восточной Европы под властью золотоордынских ханов. М., Изд-во МГУ.
Цалкин В. И. 1956. Материалы для истории скотоводства и охоты в древней Руси. «Материалы и исследования по археологии СССР», № 51. М.-Л., Изд-во АН СССР.
Цалкин В. И. 1958. Фауна из раскопок памятников Среднего Поволжья. «Тр. Куйбышевской археол. экспедиции. Материалы и исследования по археологии СССР», № 61. М.-Л., Изд-во АН СССР.
Цалкин В. И. 1960. Изменчивость метаподий и ее значение для изучения крупного рогатого скота древности. «Бюл. МОИП», отд. биол., т. LXV, вып. 1.
Цалкин В. И. 1961. Изменчивость метаподий у овец. «Бюл. МОИП», отд. биол., т. LXVI, вып. 5.
Цалкин В. И. 1964. Домашние животные Восточной Европы в раннем железном веке. «Бюл. МОИП», отд. биол., т. XIX, № 3.
Habermehl K. H. 1961. Die Altersbestimmung bei Haustieren, Pelztieren und beim jagdbaren Wild. Paul Parej. Berlin und Hamburg.
Zietzschmann О., Кrölling O. 1955. Lehrbuch der Entwicklungsgeschichte der Haustiere. Berlin.
*) В оригинале признаки древнерусской лесной и золотоордынской на стр. 123, болгарской и M. diff — на 124. OCR
Написать нам: halgar@xlegio.ru